




| EXPRESIÓN GÉNICA. SÍNTESIS DE PROTEÍNAS |
|---|
 |
| EL ADN COMO PORTADOR DE LA INFORMACIÓN GENÉTICA |
Genética → parte de la Biología que se ocupa del estudio de la herencia biológica, intentando explicar los mecanismos y circunstancias mediante los cuales se rige la transmisión de los caracteres de generación en generación. La Genética molecular estudia estos procesos desde un punto de vista químico.
• Mendel (1822-1884)
Para MENDEL los genes eran considerados como factores hereditarios que determinaban las características ext. de los seres vivos. En su época se ignoraba su composición química o su localización. Para poder referirse a ellos fueron denominados mediante letras. Así, en los guisantes, el gen A determina que las semillas sean de color amarillo y el gen a hace que sean verdes. Pero nadie sabía qué era lo que hacía que los guisantes fueran verdes o amarillos ni cómo lo hacía. Esto es, no se sabía la naturaleza de los factores hereditarios ni cuál era su mecanismo de actuación.
• Los experimentos de Griffit
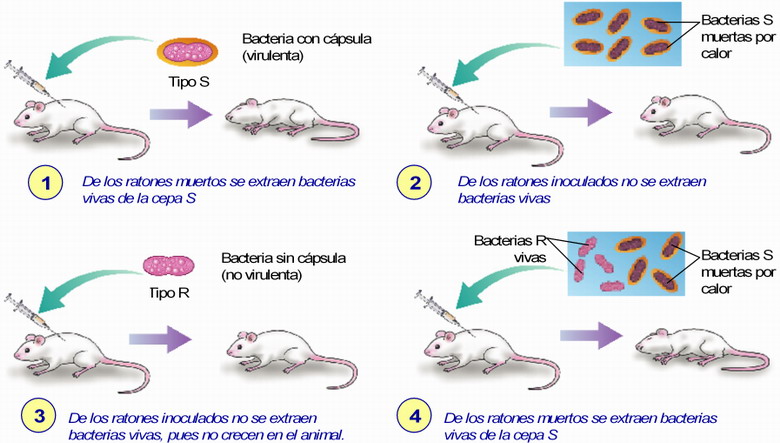
La bacteria Diplococcus pneumoniae es un pneumococo, una bacteria causante de enfermedades. Existen dos cepas, la S (Smooth = lisa), virulenta, y la R (Rough = rugosa), no virulenta. Las bacterias S, vivas, producen la muerte en los ratones, pero no la producen si están muertas. Las segundas no son capaces de desarrollar la enfermedad. En 1928 Griffith realizó con estas bacterias una serie de experiencias.
• Los experimentos de Avery (1944)
AVERY, MCLEOD y MCCARTY se propusieron encontrar cuál era el componente que transmitía el carácter heredable, y llegaron a la conclusión de que era el ADN de las bacterias S muertas por el calor el que transformaba las bacterias R en S. Demostraron así que el ADN era la molécula que contenía la información necesaria para que las bacterias S fueran virulentas y que, a pesar de estar muertas, su ADN no estaba destruido y podía pasar al medio, y de aquí a las bacterias de cepa R, integrándose en el genoma de éstas y transformándolas en virulentas.
 |
• Los experimentos de Hershey y Chase (1952)
Trabajaron con el bacteriófago T2. Introdujeron el isótopo radiactivo S35 en su cápsida proteica y el isótopo radiactivo P32 en su ADN. Infectaron un cultivo de Escherichia coli. Separadas lasa cápsidas, vieron que el S35 quedaba en las cápsulas vacías, y que el P32 quedaba en las bacterias y en el int. de los nuevos virus descendienes. Así, demostraron que era el ADN y no las proteínas, la moll. portadora de la información genética.
| Ver la REPLICACIÓN DEL ADN. Ver la EXPRESIÓN DEL MENSAJE GENÉTICO. CÓDIGO GENÉTICO. |
| TRANSCRIPCIÓN del MENSAJE GENÉTICO |
|---|
TRANSCRIPCIÓN DE LA INFORMACIÓN DEL ADN EN PROCARIOTAS
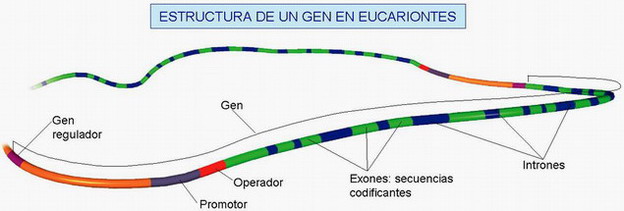
| ESTRUCTURA O REGIONES DE UN GEN (ver fig.) |
| Región promotora = promotor (P) Porción del ADN situada al principio del gen y que, sin codificar ningún aa, sirve para que las enzimas que realizan la transcripción reconozcan el principio del gen. |
| Región codificadora (C) Es la parte del gen que contiene la información para la síntesis de la proteína. En los eucariontes, existen frag. de ADN que no contienen información → intrones, y frag. que sí contienen información exones. Es preciso un proceso de →maduración que elimine los intrones y empalme los exones. Considerando la hebra 5'→ 3', el principio de esta región viene marcado por la secuencia ATG, y el final por una de estos tripletes: TAA, TAG, TGA (→ de paro, sin sentido o stop). |
| Región terminadora = terminador (T) → marca el final del gen. |
 |
| |
| TRANSCRIPCIÓN EN BACTERIAS: SÍNTESIS DE ARN |
| 1. Iniciación La ARN-polimerasa se asocia con el cofactor sigma (s), lo que le permite asociarse al promotor del gen, y polimerizar el ARN (después el cofactor s se separa). En el promotor se han encontrado dos secuencias de consenso: TTGACA, TATAAT, y secuencias intensificadoras de la transcripción. |
| 2. Elongación = alargamiento (40 nucleótidos / sg) |
| 3. Finalización Se produce al llegar a una secuencia rica en G y C, que posibilita la autocomplementariedad de la cola del ARN → un bucle final que provoca su separación del ADN. Dos variantes: una con el cofactor s y otra sin él. |
| 4. Maduración → el ARNm no tiene. El ARNt o el ARNr sí. |
| Transcripición y traducción de un gen en procariotas: 1.ADN. 2.ARNm. 3 y 4.Subunidades mayor y menor del ribosoma. 5.Proteína. a)Extremo 5´. b)Extremo 3´. c)Extremo carboxilo. d)Extremo amino. |
TRANSCRIPCIÓN DE LA INFORMACIÓN DEL ADN EN EUCARIOTAS
El ADN se encuentra en el núcleo celular y la síntesis de proteínas tiene lugar en el hialoplasma. Por ello, la información contenida en la estructura primaria del ADN debe transcribirse a una molécula del ARNm (ver fig.). También se sintetizan en el núcleo el ARNr y el ARNt, necesarios para la síntesis proteica.
| TRANSCRIPCIÓN EN EUCARIONTES: SÍNTESIS DEL ARNm |
| 1. Iniciación (ver fig.) La ARN-polimerasa II se fija en el promotor, que contiene dos señales de iniciación → secuencias de consenso: CAAT y TATA. Comienza la síntesis del ARN. |
| 2. Elongación = alargamiento (ver fig.) La síntesis de la cadena continúa en dirección 5' → 3'. Después de 30 nucleótidos se le añade al ARN una cabeza (= caperuza = líder) de metil-GTP en el extremo 5', que parece tener una función protectora para que las enzimas exonucleasas que destruyen los ARN no lo ataquen. |
| 3. Finalización (ver fig.) Cuando la ARN-polimerasa llega a la región terminadora del gen, donde está la secuencia TTATTT, finaliza la síntesis del ARNm. Entonces, una poli-A-polimerasa añade al extremo final 3´unos 200 ribonucleótidos de A → cola de poli-A, y el transcrito primario (→ pre-ARNm = ARNm precursor = ARN heterogéneo nuclear, ARNhn), se libera. |
| Maduración (ver fig.) El ARNm precursor contiene tanto exones como intrones. Es, pues, un ARNm no apto para que la información que contiene sea traducida. En la maduración, un sistema enzimático (→ ribonucleoproteína pequeña nuclear, RNPpn = complejo proteína-ARNpn) reconoce, corta y retira los intrones, y las ARN-ligasas unen los exones, formándose el ARNm maduro (ver fig.). |
Las diferencias con la transcripción en los procacariotas son las siguientes:
• En los eucariotas se produce en el núcleo. El ARNm maduro o transcrito pasa al hialoplasma donde su información servirá para la síntesis de una proteína: la información que se encuentra en forma de una cadena de nucleótidos se traducirá a una cadena de aa.
• Los procariotas sólo poseen un tipo de ARN-polimerasa. En los eucariotas hay tres:
- ARN-polimerasa I → transcribe la mayoría de los genes del ARNr.
- ARN-polimerasa II → transcribe los genes estructurales = cistrones.
- ARN-polimerasa III → transcribe los genes del ARNt y un tipo de ARNr.
• En los procariotas el ARNm no tiene ni caperuza ni cola, y no tiene intrones (por que no requiere de un mecanismo de maduración. Al mismo tiempo que el ARNm se transcribe se está ya traduciendo. En las eucariotas, hay intrones y exones (→ ARNm primario); después de la maduración sólo quedan los exones (→ ARNm maduro).
• En los procariotas los genes son policistrónicos: el ARNm contiene información para varias proteínas.
| Transcripición y traducción de un gen en eucariotas: 1.ADN. 2. ARN-polimerasa. 3.ARNmp. 4.ARN. 5.ARNm maduro. 5.Proteína. 7.Ribosoma. |
| |
| TRADUCCIÓN. SÍNTESIS DE PROTEÍNAS |
|---|
Consiste en la síntesis de una proteína a partir de la información contenida en el ARNm. Se produce en en los ribosomas del hialoplasma.
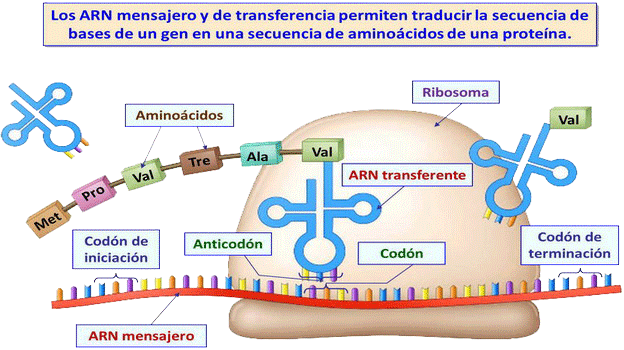 |
Etapas (ver fig. 1 y fig. 2; fig. 1 y fig. 2):
| 1. Activación de los aminoácidos (ver fig.) La formación del enlace peptídico es un proceso endergónico. Para que pueda realizarse, los aa deben de ser activados por la aminoacil-ARNt-sintetasa (una para cada aa), que une cada aa a su ARNt corresondiente (→ aminoacil-ARNt): aminoacil-ARNt-sintetasa aa + ARNt1 + ATP -----→ aminoacil-ARNt1 + AMP + PPi Cada aminoacil-ARNt tiene un triplete → anticodón, complementario de los correspondientes codones o tripletas del ARNm. Cada aa se une, por lo tanto, a un ARNt específico, que será aquel que lleve el anticodón correspondiente. |
| 2. Iniciación de la síntesis (a) En las bacterias La secuencia inicial o región líder del ARNm, que no se traduce, se une a la subunidad menor de los ribosomas. El aminoacetil-ARNt asocia su anticodón al codón del ARNm. Ahora se une la subunidad ribosómica mayor, formándose el complejo ribosomal o complejo activo, que posee dos sitios de unión: - Centro peptidil o centro P → donde se sitúa el primer aminoacil-ARNt. - Centro aceptor de nuevos aminoacil-ARNt (= región aminoacil = centro A). El primer triplete que se traduce es el AUG (→ formilmetionina), que, posteriorm., puede ser retirado. (b) En eucariotas El ARNm primario es sintetizado en el núcleo y antes da salir experimienta una maduración → ARNm maduro. Éste lleva en el extremo 5´ una caperuza (de metil-guanosin-trifosfato), que permite su identificación por los ribosomas; a cont. está la región líder, y el triple AUG (→ metionina), que, frec., es retirado. La subunidad pequeña del ribosoma se une a la región líder del ARNm, y éste se desplaza hasta que al llegar al codón AUG, que codifica el principio de la proteína. Se gasta GTP. |
| 3. Elongación de la cadena polipeptídica - El complejo ARNt-aminoácido 2 (ARNt-aa2) se sitúa enfrente del codón correspondiente en la región aceptora (centro A). - Se forma el enlace peptídico entre la metionina y el 2º aa (aa2), catalizado por la peptidil-transferasa. El centro P queda ocupada por un ARNt sin aa (que sale del ribosoma). - Se produce entonces la translocación ribosomal → el ARNm se traslada como la cinta de una máquina de escribir y el complejo ARNt-aa2-met queda situado en la región peptidil (centro P) del ribosoma y la posición aminoacil queda libre para la entrada del complejo ARNt-aa3. El ARNt de la metionina se libera. Así se van a ir añadiendo el resto de los aa que constituyen la proteína hasta llegar al codón de finalización. Se gasta GTP. |
| 4. Finalización de la síntesis Cuando el ribosoma llega al codón de finalización (uno de los codones sin sentido: UAA, UAG, UGA), la cadena polipeptídica se libera y las subunidades del ribosoma se disocian y se separan del ARNm. La estructura terciaria y cuaternaria de las proteínas se va adquiriendo según estas se van sintetizando. Una proteína puede estar constituida por una cadena polipeptídica o por varias subunidades. |
En resumen, cada codón del ARNm se le acopla un anticodón de ARNt-aa, que a su vez lleva "colgado" un aa específico. Al avanzar el ribosoma sobre el ARNm va reconociendo (traduciendo) los distintos codones a los que su unen los ARNt-aa por sus correspondientes anticodones complementarios. Los aa quedan juntos y se unen por enlace peptídico, y el ARNt se separa del ARNm y de su aa alejándose del ribosoma. El proceso se repite hasta quedar formada la proteína.
Varios ribosomas (→ polirribosoma = polisoma) de 4 a 6, a veces incluso 100, pueden estar traduciendo al mismo tiempo una cadena de ARNm.
| Ver la REGULACIÓN DE LA EXPRESIÓN GÉNICA. HIPÓTESIS DEL OPERÓN. |




